Drukuj
Jak powstało życie?
publikacja 24.10.2006 15:49
Nagromadzenie danych to nie jest jeszcze nauka. Galileusz .:::::.
WSTĘP
Prawdziwy naukowiec cechuje się tym, że zawsze szuka związków przyczynowo - skutkowych w zjawiskach, które postrzega. Widząc na przykład afrykańskiego lamparta i południowoamerykańskiego jaguara, zauważy ogromne podobieństwo między nimi. Postawi więc hipotezę o przyczynie tego podobieństwa, którą jest domniemany wspólny przodek obydwu gatunków. Żył on później, niż wspólny przodek tych kotów i na przykład wilka, i dlatego są one bardziej podobne do siebie niż do swojego kolegi z rodziny psowatych. Miały więcej czasu na zmiany. Mimo faktu, że obydwa wspomniane koty są znakomitymi pływakami, trudno byłoby przypuszczać, że któryś z nich pokonał dystans dzielący obydwa kontynenty wpław. Naukowiec wysunie więc hipotezę, według której dystans ten został pokonany drogą lądową przez Azję i Cieśninę Beringa. Hipotezy mogą później znaleźć potwierdzenie w badaniach paleontologicznych.
Istnieją także inne hipotezy: podobieństwo przypadkowe lub konwergencja, ale są one słabe, bo podobieństwa między wymienionymi kotami są zbyt szczegółowe, w zbyt wielu elementach budowy, a także niepowierzchowne.
Jeśli założymy, że przodkiem wszystkich żyjących obecnie organizmów była jedna prakomórka, istniejąca około 3,8 mld lat temu, to pojawia się poważny problem. Ona się przecież nie zachowała. Więc jeśli się nie zachowała, to problem wydaje się być nierowiązywalny. To tak jakby na podstawie wyglądu i położenia śniegu leżącego u stóp góry, po której stoku zeszła lawina, „odtworzyć” miejsce, z którego zsunął się kamień - sprawca lawiny oraz jego kształt, rozmiar i masę.
Pierwsza żywa prakomórka z pewnością istniała. To był taki mały kamyk, który uruchomił lawinę doprowadzającą po 3,8 mld lat do takiej różnorodności form organizmów żywych na Ziemi. Ale co możemy o niej powiedzieć?
Zagadka powstania życia nie jest na szczęście tak trudna, jak zagadka lawiny zsuwającej się z góry – przykład analogii pozornej. Prakomórka przecież dzieliła się, więc ta lawina różnorodności biologicznej, którą uruchomiła miała „coś z niej”. Różnica nie jest więc tu tak drastyczna, jak między kamieniem a śniegiem.
Problem początków życia rzuciłyby z pewnością na kolana nawet samego Sherlocka Holmesa, ale nie jest skrajnie beznadziejny.
Prakomórka - „przestępca”, dzieląc się zostawiał ślady. Nawet ten, który żył 3,8 mld lat temu... Potomkowie są przecież podobni do przodków.
STRATEGIA POBIERANIA ŚLADÓW
Pozostaje więc pytanie, pod jakim względem i na ile są podobni. Prawie 4 mld lat to szmat czasu, miliony pokoleń i wiele „okazji” do zmian, różnicowania się, z czym kojarzy nam się zacieranie śladów. Zacieranie przez czas. A poza tym skąd będziemy wiedzieć które cechy, „ślady” mogły być charakterystyczne dla prakomórki, a którymi nie warto się zajmować, bo przyszły później? Spójrzmy na poniższy rysunek:
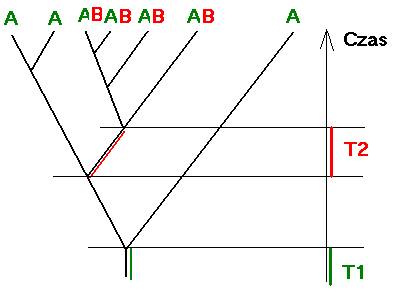
Cecha A obserwowana jest u wszystkich organizmów, widocznych na rysunku drzewa ewolucyjnego. Cecha B obecna jest tylko na zakończeniach jednego z odgałęzień. Pozwala to przypuszczać, że cecha A powstała wcześniej niż cecha B; w interwale czasowym T1.
Cechy prakomórki powstały najwcześniej, bo przed nią nic nie było. A więc musimy szukać jej śladów w cechach wspólnych dla całego świata ożywionego.
Czy takie są ? Są szanse, że odpowiedź na to pytanie jest pozytywna. Teoretycznie możliwy jest zanik tych najbardziej prymitywnych cech na każdej z linii ewolucyjnych, prowadzących od prakomórki do współczesnych organizmów. Co może jednak stanąć na przeszkodzie?
Wyobraźmy sobie budowlę postawioną 100 lat temu. Możliwe jest dobudowanie do niej wież, wymiana okien, remont wnętrz. Ale najtrudniej byłoby zmienić fundamenty, czyli to co powstało najwcześniej i od czego uzależniona w sposób podstawowy jest cała budowla. Jest rzeczą niemal pewną, że „ruszenie” fundamentów (ich zmiana) spowoduje poważne uszkodzenia budowli.
A więc może w obecnych komórkach istnieją takie kopiowane i nienaruszane fundamenty prakomórki?
JEDNOŚĆ ŚWIATA OŻYWIONEGO
Strukturalna: to, co rzuca się w oczy przy ogólnym oglądzie budowy organizmów, to ich budowa komórkowa. A każda komórka ma cytoplazmę, materiał genetyczny i okalającą je błonę komórkową.
Biochemiczna:
- Podstawowym składnikiem błon komórkowych są fosfolipidy.
- Wszystkie organizmy są w stanie produkować glukozę (z udziałem obcego lub własnego źródła energii), metabolizować glukozę w procesie glikolizy. Mają też cykl pentozofosforanów metabolizujący glukozę i wytwarzający rybozę (istotny składnik DNA).
- Wszystkie białka organizmów żywych, to różne kombinacje tego samego zestawu 20 aminokwasów.
- Prawie wszystkie reakcje zachodzą w organizmach żywych szybko, dzięki udziałowi białkowych enzymów.
- Materiał genetyczny wszystkich organizmów żywych, to DNA, złożony zawsze z fosforanów deoksyrybonukleozydów A,T,G,C, przepisywany na mniej trwały RNA, złożony zawsze z fosforanów rybonukleozydów A,U,G,C.
Genetyczna:
- Wszystkie organizmy żywe mają transkrypcję (przepisywanie informacji genetyczne z DNA na mniej trwały nośnik – RNA).
- Wszystkie organizmy żywe mają translację - tłumaczenie informacji genetycznej na język białek w rybosomach, których kluczowym składnikiem jest zawsze rRNA. Do procesu tego wymagane są uniwersalne cząstki tłumaczące – tRNA.
- Wszystkie organizmy żywe mają taki sam kod genetyczny, przypisujący jednej z (64 - 3 = 61) trójkowych kombinacji zasad azotowych, dany aminokwas.
ŚCIANA PROBLEMÓW
Musimy wziąć pod uwagę istotny fakt mówiący, że około 3,8 mld lat temu na Ziemi nie było nikogo, kto mógłby stworzyć pierwszą prakomórkę. Jednym z filarów nowożytnej nauki jest materializm będący przekonaniem, że wszystko da się w nauce wyjaśnić i wyprowadzić bez ingerencji żadnych tajemniczych, nieuchwytnych, duchowych czynników. Krótko mówiąc, dla naukowca, materia, będąca budulcem pierwszej prakomórki, musiała zorganizować się sama, bez niczyjej, tajemniczej pomocy.
Samoorganizacja materii w żywą komórkę nie jest sprzeczna z prawami fizyki i chemii. Nikt takiego dowodu nie przedstawił. Naukowcy mają więc „przepustkę” do poszukiwania etapów tego procesu. Do tego muszą oni jednak wiedzieć czym jest życie, bo aby wiedzieć jak coś może powstać, trzeba też wiedzieć co ma powstać. Na pytanie to odpowiada artykuł Czym jest życie?
Z tego, co do tej pory wiemy, wynika, że prakomórka musiała mieć błonę komórkową, cykl metaboliczny dostarczający składników budulcowych, enzymy przyspieszające przebiegi reakcji metabolicznych i materiał genetyczny.
Niezwykle problematyczna pozostaje niestety synteza niektórych istotnych składników budulcowych, sposób i kolejność ich organizacji.
Oto zestaw problemów biogenetycznych, z którymi przyjdzie nam się teraz zmierzyć:
Problem 1 Jak przebiegła synteza podstawowych cegieł budulcowych żywych komórek: aminokwasów, glukozy, rybozy, zasad azotowych będących składnikami RNA i DNA?
Problem 2 Co stanowiło składnik otoczki prakomórek, skoro synteza fosfolipidów jest skomplikowana i wymaga zaawansowanego aparatu enzymatycznego?
Problem 3 Skąd prakomórka czerpała energię i jaki był jej metabolizm?
Problem 4 DNA koduje informację genetyczną, ale do swej syntezy i replikacji wymaga białek. Białka katalizują też reakcje metaboliczne, a niektóre są budulcem komórki. Ale informacja o sekwencji białka jest zapisana w DNA. Co było pierwszym materiałem genetycznym i czym były pierwsze enzymy, skoro białka potrzebują DNA do powstania i odwrotnie?
Problem 5 Jak przebiegała replikacja pierwszych kwasów nukleinowych?
Problem 6 Jakie były pierwsze nici informacji genetycznej. W wyniku braku wspomagania przez białka, słabej stabilizacji wiązań między zasadami azotowymi, nieumiejętności rozpoznania początku odczytu, jakie musiały być pierwsze kodony?
Problem 7 Dlaczego w komórkach obserwuje się swoiste złamanie symetrii i w ich skład wchodzą właściwie zawsze D-cukry i L-aminokwasy (a nie na przykład L-cukry i D-aminokwasy)?
Problem 8 Jak powstał tRNA i pierwsze, prymitywne rybosomy do syntezy białek?
Problem 9 Jak powstał uniwersalny kod genetyczny?
Problem 10 Jak powstały chromosomy, zawierające ułożone na niciach DNA geny (zamiast istnienia każdego z osobna na oddzielnym kawałku DNA)?
Problem 11 Gdzie mogła powstać pierwsza prakomórka, aby przy jej stopniowym tworzeniu się składniki nie rozdyfundowywały?
LABORANTKA ZIEMIA
W zamierzchłych czasach – około 3,8 mld lat temu, atmosfera ziemska miała charakter redukujący. Geolodzy wiedzą to stąd, że skały pochodzące z tamtego okresu są słabo utlenione. W atmosferze tej znajdowały się prawie na pewno w dużych ilościach takie gazy jak: metan, amoniak, wodór, para wodna, formaldehyd.
W swoim słynnym doświadczeniu z 1953 r. S. Miller z Uniwersytetu w Chicago odtworzył przypuszczalne warunki panujące w ziemskiej atmosferze 3,8 mld lat temu. Do kolby wpuścił mieszaninę gazów: metanu, wodoru, amoniaku i pary wodnej i działał na nią wyładowaniami elektrycznymi, mającymi symulować wyładowania atmosferyczne.
W produktach reakcji odkrył duże ilości aminokwasów: glicyny, alaniny, kwasu asparaginowego, waliny. W zauważalnych ilościach produkowane były też: kwas glutaminowy, leucyna, izoleucyna, seryna i treonina. W mieszaninie poreakcyjnej stwierdzono też znikome ilości zasad azotowych: pirymidynowych i purynowych. Nie było w niej natomiast żadnych lipidów.
W reakcji chemicznej, tzw. reakcji formozowej, której substratem jest tylko formaldehyd, powstają monocukry w postaci racematów (czyli zarówno D jak i L).
n HCHO → CH2OH(CHOH)(n+2)CHO n = 0,1,2,3,4
Reakcja przebiega szybciej w obecności monocukrów, więc produkty reakcji powodują, że z czasem staje się ona coraz wydajniejsza. Wydajność produkcji rybozy w tej reakcji wynosi około 1%. Oto w miarę zadowalająca odpowiedź na postawiony problem 1.
Na skałach w tych odległych czasach istniały z pewnością wgłębienia, dołki, małe kałuże, okresowo zalewane opadami, bogatymi w produkty reakcji gazów w ziemskiej atmosferze.
Roztwory tych związków dzięki temu nie musiały być bardzo rozcieńczone, a nawet z czasem mogły zatężać się. Jaki byłby tego mechanizm? Jeśli do szklanki z roztworem soli będziemy dodawać dalej ten roztwór uzupełniając nim wodę, która wyparowała, to będziemy z czasem otrzymywać roztwór o coraz to większym stężeniu. To raczej w takich izolowanych zagłębieniach, kałużach, a nie na otwartych morzach przebiegł proces biogenezy (problem 11).
DETEKTYWI BIOGENEZY
A co z pozostałymi problemami ? Są one bardzo zawiłe i ich rozwiązanie, czy przynajmniej próby, wymagały pracy i współpracy wielu znakomitych naukowców. Oto najważniejsze wyniki ich zmagań:
a) Detektyw Oparin
Każda z istniejących obecnie komórek otoczona jest podwójną błoną lipidową - plazmalemmą. Stanowi ona granicę pomiędzy zawartością komórki a środowiskiem i zapobiega dyfuzji jej składników.
Oparin zauważył, że różne polipeptydy i polisacharydy, rozpuszczone w wodzie samorzutnie formują pęcherzyki. Nazwał je koacerwatami. Pierwotna plazmalemma mogła stworzyć się więc spontanicznie, ze związków których jednak dokładnej budowy chemicznej nie poznamy nigdy. Obecne błony lipidowe są więc pozostałością po niej, choć raczej niewiele przypominają ją z chemicznego punktu widzenia. Wnioskując z doświadczenia S. Millera lipidy musiały pojawić się później, więc początkowo musiały być zastępowane przez inne związki chemiczne.
Koacerwaty Oparina potrafią gromadzić w swoim wnętrzu enzymy, które są w stanie dokonywać złożonych przemian chemicznych. Problem z koacerwatami wynika jednak stąd, że nie posiadają one same właściwości katalitycznych. Są tylko w stanie pobierać enzymy, zgodnie z zasadą rozdziału substancji pomiędzy dwie fazy (koacerwat i środowisko) tzn. enzym zgromadzi się w większym stężeniu w tej fazie w jakiej jest lepiej rozpuszczalny.
Trudno jednak zakładać spontaniczne formowanie enzymów - skomplikowanych białek w środowisku, w którym zanurzone były koacerwaty. Są to związki, które w wodzie szybko hydrolizują. Nie potrafią się również same kopiować, a szansa powstania przypadkowo m peptydów o takiej samej sekwencji n - aminokwasowej wynosi: 1 / (20^(nxm)) i bardzo szybko maleje ze wzrostem m i n.
Koacerwaty Oparina stanowiły pewien krok naprzód, gdyż pokazał on, że polisacharydy potrafią spontanicznie formować pęcherzyki komórkopodobne i wejść w skład ich otoczki zamiast lipidów (problem 2).
b) Detektyw Cairns-Smith
Kryształy glinokrzemianów, z rozmieszczonymi wewnątrz ich sieci krystalicznej jonami metali, mogły pełnić funkcje katalizatorów przemian chemicznych. Miały one z pewnością niską wydajność. Pozostaje to na razie w sferze hipotez, gdyż nikt jeszcze doświadczalnie nie udowodnił katalitycznych zdolności kryształów gliny, a żadnych pozostałości glinokrzemianowych z tamtego okresu obecnie w komórkach na razie nie stwierdzono. Można by więc zapytać: niewinność to czy "przestępstwo" doskonałe?
Pierwotne polimery, których syntezę przeprowadzały glinokrzemiany mogły według szkockiego chemika - Cairns-Smitha składać się tylko z cukrów i reszt fosforanowych. Taki polimer fosfocukrowy stanowi dzisiaj rdzeń kwasów nukleinowych (zob. rys.3). Prakałuża obfitowała w formaldehyd, ten zaś bardzo łatwo kondensuje z wytworzeniem monocukrów, zgodnie ze wspomnianą już reakcją formozową. Jony fosforanowe w wodzie morskiej pochodziły najprawdopodobniej z rozpuszczanych przez nią stopniowo skał.
Gunter Wachtershauser wysunął niedawno hipotezę mówiącą, że wiele reakcji chemicznych, istotnych do procesu biogenezy mogło zachodzić na powierzchni kryształów pirytu (dwusiarczku żelaza II). Powierzchnia ta naładowana jest dodatnio, więc mogła przyciągać jony fosforanowe i wspomagać też katalizę polimerów fosfocukrowych. Reakcje przeprowadzane na powierzchni były bardziej wydajne, bo łatwiej monomerom spotkać się w powierzchni niż w przestrzeni. Niewykluczone, że struktura powierzchni pirytu mogła wymuszać reakcje reszt fosforanowych z grupami 5’ i 3’ rybozy.
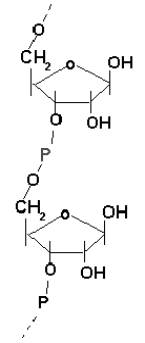
Rys.2. Wzór strukturalny kwasu polifosforybozowego, który mógł istnieć przed powstaniem RNA.
Z czasem powstawały bardziej skomplikowane cząsteczki, np. RNA. Niektóre z nich zaczynały przeprowadzać reakcje metaboliczne z dużo większą szybkością. Zjawisko przejęcia władzy od kryształów przez RNA, zostało nazwane przez Cairns Smitha: genetic takeover (genetyczne przejęcie władzy).
A skąd przekonanie, że pierwszym kwasem nukleinowym był RNA ? Po pierwsze dlatego, że w reakcji formozowej powstaje spontanicznie ryboza, a nie deoksyryboza. A po drugie... I tu zatrzymamy się na dłużej.
Wiemy, że we współczesnych komórkach DNA pełni funkcję przechowywania informacji genetycznej. Funkcję katalityczną pełnią białkowe enzymy. Można się jednak doszukać w komórkach śladów pierwotnego, katalitycznego RNA. Oto niektóre z nich:
Selfsplicing
Jest to proces występujący m.in. w cząsteczkach rRNA pierwotniaka Tetrahymena oraz w wielu mRNA genów mitochondrialnych u drożdży. Polega na wycięciu intronu z RNA, katalizowanym przez niego samego, bez udziału białek.
RNaza P
Jest to enzym służący u Procaryota do obróbki powstałego po transkrypcji tRNA. Enzym zawiera białko i RNA. Właściwości katalityczne ma jednak tylko ten drugi składnik, co pozwala sądzić, że był ważniejszy i wcześniejszy w ewolucji. Białko stabilizuje tylko strukturę przestrzenną RNA.
Rybosomy
Procaryota - 3 rodzaje rRNA, 52 białka
Eucaryota - 4 rodzaje rRNA, 82 białka
Tylko rRNA w rybosomach pełni funkcje katalityczne, natomiast ani jedno białko lub jakakolwiek ich grupa nie ma takich właściwości. Służą one do stabilizowania struktury trzeciorzędowej rRNA i zwiększenia przez to jego wydajności katalitycznej.
Widzimy więc, że RNA o właściwościach katalitycznych - rybozymy przetrwały w szczątkowej formie do dziś. Istnieją one jednak najczęściej pod zwałami białek, których lawina stoczyła się na nie w trakcie ewolucji.
Naukowcom udało się też sztucznie wytworzyć kilka rybozymów katalizujących reakcje metaboliczne, takie jak: biotynylacja, aminoacylacja, dodanie reszty fosforanowej, ligacja cząstek RNA.
RNA to bardzo ciekawy związek chemiczny. Jego cechy stanowią syntezę najważniejszych cech białka i DNA. Białka są świetnymi katalizatorami, ale nie potrafią replikować się, a do zwiększenia liczby swych kopii potrzebują zapisu genetycznego w DNA i procesu translacji. DNA nie przeprowadzi żadnej reakcji chemicznej, gdyż ma formę podwójnej helisy i wszystkie zasady azotowe, które mogłyby być katalitycznie aktywne są schowane wewnątrz niej. DNA potrzebuje więc białek, a więc nie mógł być pierwszy.
RNA jest już natomiast takim małym „kwaziorganizmem” bez metabolizmu. Jego sekwencja stanowi genotyp - całkowity zapis genetyczny, struktura przestrzenną zaś – fenotyp, czyli zestaw cech struktury przestrzennej, determinowany przez zapis genetyczny, dzięki któremu może on stać się enzymem.
Oto rozwiązanie problemu 4. Na początku funkcje enzymów i DNA pełniły dwoiste kwasy rybonukleinowe - RNA.
Jak jednak powstały cząsteczki RNA z łańcucha polifosforybozowego?
Stawiam tutaj hipotezę o powstaniu zasad azotowych RNA, wykorzystując wiedzę biochemiczną o współcześnie istniejącym metabolizmie.
Biosynteza puryn odbywa się w komórkach z udziałem glicyny, formaldehydu i amoniaku (specyficznie związanych). Biosynteza pirymidyn wymaga natomiast kwasu asparaginowego, amoniaku i dwutlenku węgla. Wszystkie te substancje występowały już w prakałuży i praatmosferze. Zdumiewająca zgodność !
Związek z rys.2 mógł więc mieć najpierw doczepiany w miejsce grup 1’OH amoniak, potem aminokwasy, co zwiększało jego zdolności katalityczne. Teraz mógł katalizować dalsze dobudowywanie reszt HCHO i NH3 na innych łańcuchach, aż do całkowitego zsyntetyzowania zasad azotowych (zob. rys.3)
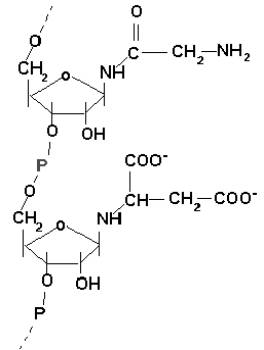
Rys.3. Wzór strukturalny domniemanego związku po dołączeniu do kwasu polifosforybozowego reszt amoniaku, glicyny i kw.asparaginowego.
c) Detektyw Eigen
Manfred Eigen skupił swą uwagę na cząsteczkach zdolnych do replikacji. Jego eksperyment z replikazą RNA, pochodzącą z wirusa o nazwie Qbeta pokazał, że replikowane przez nią cząsteczki RNA „walczą” o byt, a zmagania te zawsze doprowadzają do wyselekcjonowania cząsteczki, która replikuje się najwydajniej w środowisku, które stanowi określone stężenie soli i temperatura.
Eksperyment jest powtarzalny. Problemem jest fakt, że replikaza jest białkiem, które miało przecież w ewolucji powstać później. Orgel wykazał jednak, że cząsteczki RNA są zdolne do autoreplikacji. Trzeba im tylko dostarczyć rybonukleotydów i kationów cynku.
J. Maynard Smith i E. Szathmary udowadniają, że przy założeniu, że szybkość replikacji cząsteczek RNA na dwie wolne nici jest znacznie większa niż szybkość dysocjacji podwójnej helisy RNA, powstałej w replikacji, możliwa jest stabilna koegzystencja różnych cząstek RNA. Charakteryzuje je wtedy wzrost paraboliczny. Liczy się nie tylko szybkość produkcji kopii, ale także szybkość oddysocjowywania wolnych, pojedynczych nici do dalszej syntezy.
Wzrost opisuje równanie: dA/dt = kA^p , gdzie p<1. W takim przypadku cząsteczki RNA nie „wygryzają się” wzajemnie w taki sposób, że zostanie tylko jedna.
Wzrost, który zależy tylko od szybkości produkcji kopii opisywany wzorem:
dA/dt = kA^p, (p=1) to wzrost ekspotencjalny charakterystyczny dla teoretycznego, nieograniczonego wzrostu kolonii komórek. Tam czynnika dysocjacji nie ma, bo one nie są ze sobą „sklejone” wiązaniami chemicznymi.
Cząsteczki RNA potrafią mieć własności katalityczne. Może więc potrafią wzajemnie wspomagać się w replikacji? Myśl ta naprowadziła Manfreda Eigena na ideę hipercykli (zob. rys.4).
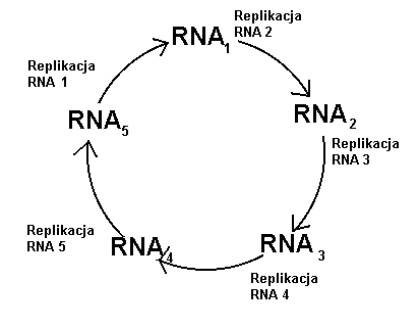
Rys.4. Schemat hipercyklu.
System RNA wspierających się w kopiowaniu, jest stabilny i samopodtrzymujący się. Żadna pojedyncza cząsteczka nie jest w stanie zdominować reszty. W interesie każdej leży kooperacja.
Tutaj przyrost stężenia danej cząstki RNA będzie zależał „silniej” niż tylko od jej stężenia, bo będzie ona w swym kopiowaniu wspomagana.
Czyli dA/dt = kA^p, p>1. Jest to tzw. wzrost hiperboliczny.
Wygrana hipercyklu zależy nie tylko od szybkości replikacji, ale także od początkowego stężenia jego składowych.
Dwóch naukowców z USA N. Hud i F. Anet wysunęło ostatnio hipotezę mówiącą, że replikacja RNA musiała być wpierana przez związki porfirynopodobne, które interkalowały między zasady RNA, usztywniały je i przyciągały i stabilizowały zasady komplementarne. Hipoteza wyciągnięta była na podstawie danych empirycznych, które nie potwierdzały satysfakcjonująco zdolności replikacji RNA pod nieobecność enzymów białkowych. Być może tutaj bierą swój początek porfiryny, występujace we współczesnych organizmach. Na uwagę zasługuje również fakt, że w metabolizmie współczesnych organizmów przeprowadzającym rybozę w deoksyrybozę wymagana jest cyjanokobalamina – związek porfirynowy (!).
Jak długie mogły być pierwotne łańcuchy RNA? Autokataliza przeprowadzana jest z błędem około 1 nukleotyd/20. To znacznie za dużo. Z udziałem enzymów białkowych margines błędu wynosi tylko 1/1000. A więc należy przypuszczać, że replikacja wspomagana przez mniej specyficzne rybozymy miała margines błędu 1/100-200 i cząsteczki replikowane mogły mieć długości tego rzędu co dzisiejsze tRNA: 100 do 200 nukleotydów.
Po replikacji powstaje podwójna helisa RNA. Bardzo prawdopodobne, że procesy syntezy i dysocjacji były ponad 3,5 mld lat temu związane z cyklami dnia i nocy. W dzień temperatura wzrastała, dostarczając energii do rozerwania wiązań w zasadach komplementarnych (→dysocjacja). W nocy, gdy było zimniej, następowała autoreplikacja lub replikacja cząstek RNA. Oto odpowiedź na problem 5.
d) Detektyw Maynard Smith
John Maynard Smith utrzymuje, że kwasy nukleinowe, niosące już ze sobą informację genetyczną pierwotnie składały się tylko z nukleotydów guaninowych - G i cytydynowych - C. Działo się tak dlatego, że replikacja cząsteczek RNA, składającego się tylko z tych nukleotydów cechuje się największą dokładnością, czyli najmniejszą liczbą błędów. Nieprzypadkowo więc w kodzie genetycznym dwa aminokwasy powstające w największych ilościach w doświadczeniu Milllera reprezentowane są przez kodony: GGC, GGG (glicyna) i GCG, GCC (alanina). Smith założył że pierwotny aparat translacji nie był na tyle dokładny, by rozpoznać start właściwej ramki odczytu. Tak więc, aby zapobiec wytwarzaniu, w dwóch przypadkach na trzy, peptydów nonsensownych kod mógł opierać się najpierw tylko na dwóch kodonach: GGC i GCC.
Ułóżmy z nich dowolną sekwencję:
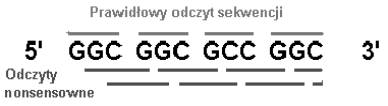
Antykodony na odpowiednich tRNA pasujące do kodonów powyżej to: 5’ GCC 3’ i 5’ GGC 3’. Przypatrując się ich sekwencji, można zauważyć ciekawą rzecz. Sekwencje antkodonowe są takie, jak sekwencje kodonów! Pierwotne mRNA i tRNA mogły więc ewoluować wspólnie od jakichś cząsteczek RNA, mających tylko 2 kodony, z których część wyspecjalizowała się w niesieniu informacji, a część w jej tłumaczeniu na język aminokwasów! Antykodony te będą parować do kodonów tylko w jednej ramce odczytu. Odczyt w ramce pierwszej na dole w rysunku powyżej zaczyna się zawsze od środkowego nukleotydu (C lub G) i kończy na G. Daje więc kodony GCG i CCG. Odczyt w ramce drugiej na dole zaczyna się zawsze od C i kończy na C lub G. Daje kodony CGG i CGC. Z wszystkimi czterema kodonami antykodony nie będą więc parować.
Ponadto, podczas replikacji powyższej sekwencji, na nici komplementarnej też powstaną kodony o rozpoznawalnym motywie 5’ G*C 3’:
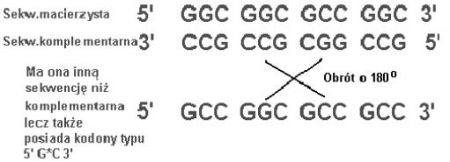
Druga nić mogła więc również służyć do kodowania jakiegoś polipeptydu, czego w przypadku DNA zazwyczaj nie obserwujemy.
Jeśli jeszcze założymy, że pierwotne nici miały symetrie środkową, czyli były tzw. palindromami i ich zakodowana sekwencja nukleotydowa była taka sama czytając od strony 5’ i 3’, np.:
5’ GGC GCC GGC GCC 3’
3’ CCG CGG CCG CGG 5’
to zauważmy, że nić po replikacji jest identyczna z nicią matrycową, a nie komplementarna. Zauważmy różnicę: nić komplementarna do sekwencji przykładowej i przypadkowo napisanej: 5’GCC TTG CCC ATG 3’ to 5’CAT GGC CAA GGC 3’ i nie jest nawet do matrycowej podobna.
Podczas replikacji palindromu otrzymujemy więc 2 nici niosące taką samą informację, a w czasach gdy same nici były „kwaziorganizmami” miało to ogromne znaczenie ewolucyjne. Najpierwotniejsze nici RNA mogły być więc palindromowymi sekwencjami złożonymi tylko z trójek GGC i GCC.
Najczęściej występujące mutacje to tranzycje G → A oraz C → U. Jeśli cząsteczki RNA mutowały z założeniem silnej presji selekcyjnej na zachowanie korzystnych przy replikacji flanek G*C to powstawały dwa następne kodony: GUC i GAC. Również nieprzypadkowo kodują one dziś odpowiednio: walinę i kwas asparaginowy - następne po glicynie i alaninie, powstające w największych ilościach aminokwasy z doświadczenia Millera. To wszystko układa się w pewną całość! I stanowi w miarę satysfakcjonującą odpowiedź na problem 6.
e) Detektyw Di Giulio
Cząsteczki transferowego RNA (tRNA) biorą udział w biosyntezie białek na matrycy mRNA. Cały proces wspomagają, stabilizują i czynią precyzyjnym rybosomy – składniki komórek zbudowane z białek i rRNA. Tak jest teraz.
A jak było kiedyś? Cząstki tRNA liczą od 75-85 zasad. Jest to niewątpliwie cecha dość pierwotna, gdy RNA nie mogły być zbyt długie, bo zbierałyby zbyt dużo mutacji. Przyjrzyjmy się więc tym cząsteczkom bliżej. Oto uproszczony schemat typowego tRNA:
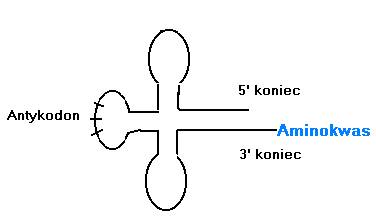
Włoski biolog teoretyk – Massimo di Giulio w swoich kilku znakomitych pracach ogłaszanych na łamach „Journal of Theoretical Biology”: 159 (1992), 171 (1994), 177 (1995), 187 (1997), 191 (1998) przyjrzał się bliżej cząsteczkom tRNA.
Przetestowana została przez niego hipoteza bezpośredniego podwojenia, jako drogi powstania cząstek tRNA. Spójrzmy na rysunek:
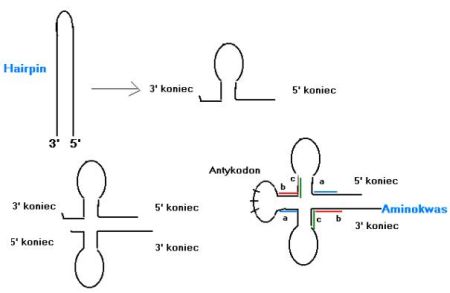
Hipoteza zaproponowana przez Hopfielda zakładała, że współczesne cząstki tRNA powstały w wyniku złożenia się 2 krótkich czątek typu hairpin (spinka do włosów). Były one stabilne, bo potrafiły zginać się w pół i parować same siebie.
Di Giulio postanowił przetestować tę hipotezę. Skoro dzisiaj tRNA powstały z 2 identycznych cząstek hairpin, to ta identyczność musi mieć jakieś ślady w dzisiejszych tRNA, bo one wolno ewoluują. Przykłady sekwencji, które powinny wykazywać podobieństwa zaznaczyłem kolorem na rysunku powyżej.
Hipoteza została przetestowana na bogatym zestawie danych i za pomocą metod statystycznych. Okazało się, że z bardzo wysokim prawdopodobieństwem podobieństwo to nie jest przypadkowe i wskazuje na bezpośrednie podwojenie.
M. Di Giulio wysunął również hipotezę mówiącą, że początkowo cząsteczki typu hairpin reagowały z aminokwasami, które stanowiły pierwsze koenzymy dla nich jako dla rybozymów w świecie bezbiałkowym. Dwie cząstki hairpin, „naładowane” różnymi aminokwasami mogły parować ze sobą jak przy utworzeniu tRNA.
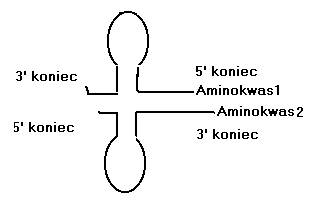
Z czasem powstawały coraz dłuższe łańcuchy polipeptydowe, podłączone do RNA (pierwsze rybonukleoproteiny). Niewykluczone, że już cząsteczki typu hairpin rozpoznawały kodony na prymitywnych mRNA za pomocą „preantykodonów” na swych zagięciach. Z czasem powstawały pełne tRNA. Zauważmy że pętli antykodonu odpowiada 3’koniec. Możliwe więc, że prymitywne enzymy, przypisujące aminokwas do RNA, nie rozpoznawały odległego antykodonu, lecz identyczną sekwencję na 3’ końcu – czyli tam gdzie aminokwas podczepiany jest do dziś, choć dzisiaj rozpoznawany antykodon jest odległy, a identyczność z 3’ końcem zatarł czas. Proces prymitywnej translacji musiał być stabilizowany, ale początkowo nie przez białka, ale przez zlepek wielu RNA, stanowiących centrum organizujące, które katalizowało powstanie wiązań peptydowych i stabilizowało wiązanie tRNA do mRNA. Francis Crick – współodkrywca struktury DNA powiedział kiedyś, że prymitywny aparat translacyjny (prarybosom) musiał opierać się tylko na RNA (rRNA). I trudno się z tym nie zgodzić, nie tylko wskutek wywodów logicznych (białek na początku być nie mogło, bo najpierw trzeba było je wytworzyć), ale także empirycznie - badając współczesne rybosomy. Tylko RNA pełni w nich funkcje katalityczne, a białka całość stabilizują i zwiększają wydajność translacji.
Oto w miarę satysfakcjonujące rozwiązanie problemu 8.
f) Detektyw Wong
J. Tzei-Fei Wong ogłosił swoją teorię ewolucji kodu genetycznego, tzw. teorię koewolucyjną w 1975 roku w swojej znakomitej pracy, którą warto tu zacytować.
WONG TZEI-FEI J. A coevolution theory of the genetic code. Proceedings of the National Academy of Sciences of the USA (1975) 72(5), 1909-1912.
Jest artykuł jest krótki, ale esencjonalny i treściwy, więc z pewnością warto doń dotrzeć. Wong roztacza tam swoją hipotezę odnośnie przypisywania trójek kodonów danym aminokwasom. Przed nim postulowano również teorię stereochemiczną (jest związek między kodonem, a aminokwasem w rodzaju ich dopasowania przestrzennego). Teoria ta nie zdobyła jednak silnego poparcia w faktach.
W latach sześćdziesiątych XX wieku Francis Crick wysunął hipotezę mówiącą, że każdy kodon przypisany jest każdemu z 20 aminokwasów (+ kodony STOP) tak jak przypadek zdarzył i nie ma tu żadnej regularności. Taki przypadkowy kod utrwalił się i dlatego idea Cricka została nazwana Hipotezą Zamrożonego Przypadku. Zamrożony przypadek (ang. frozen accident) to idea warta wyjaśnienia. Wyobraźmy sobie osiedle budowane 30 lat temu. Przed zbudowaniem bloku były do wyboru cegły i płyty betonowe. Zostały wybrane płyty betonowe i teraz po 30 latach nikt już tych płyt cegłami nie zastąpi. Trzebaby wyburzać całe osiedle, a to nie jest realne. Podobnie z kodem genetycznym w komórkach. Wybrane zostało przypadkowe przypisanie aminokwasów kodonom i teraz jest to wybór nie do odwrócenia (choć tylko przypadkowy).
Z hipotezą tą nie zgodził się J. Tzei-Fei Wong, zauważając ciekawe prawidłowości w kodonach aminokwasów i ich ścieżkach metabolicznych (rys.5).
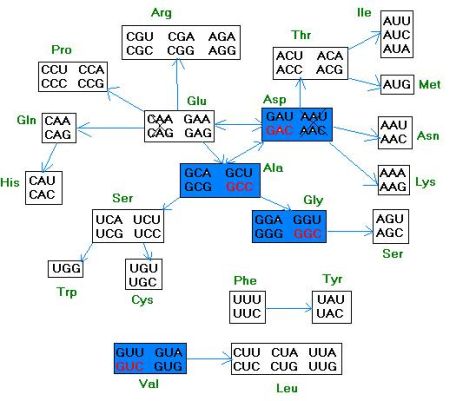
Rys.5 Schemat zależności pomiędzy ścieżkami metabolicznymi aminokwasów, a płynnymi, ciągłymi przejściami pomiędzy ich kodonami.
Na początku były kodony GGC (glicyny) i GCC (alaniny). Potem doszły kodony kwasu asparaginowego (GAC) i waliny (GUC). Należy pamiętać, że pary AU tworzą tylko dwa wiązania, a pary GC – trzy. Powstanie RNA niosących uracyl (U) i adeninę (A) mogło ułatwiać dysocjację podwójnej helisy RNA po replikacji w celu przejścia następnego cyklu replikacji. Takich cząstek było z czasem zapewne więcej.
Z czasem powstawały kodony dla treoniny, seryny, izoleucyny, leucyny i kwasu glutaminowego. Kodony stop były najprawdopodobniej 4: UAA, UAG, UGA, UGG. Tylko jeden kodon - UGG z czasem został „przypisany” tryptofanowi i jest to teraz najrzadziej występujący aminokwas w białkach.
Strzałki na powyższym rysunku wskazują na kierunki i kolejności przemian biochemicznych aminokwasów. Niesłychanie wyraźna i zdumiewająca jest regularność typu: im bliżej inny aminokwas znajduje się od danego aminokwasu, tym jego kodon wymaga mniej zmian, aby przejść w kodon danego sąsiada szlaku metabolicznego. To tak jakby za każdym razem, gdy powstawała nowa ścieżka metaboliczna (dzięki istnieniu rybozymów wspomaganych kofaktorami aminokwasowymi), prowadząca do powstania aminokwasu, przypisany zostaje mu kodon różniący się tylko jedną mutacją względem kodonu prekursora.
I tak prawie na pewno było. I tak prawie na pewno powstawał kod genetyczny.
Zauważmy również, że tak powstający kod genetyczny jest „bezpieczniejszy”. Aminokwas sąsiadujący w ścieżce metabolicznej jest z dużym prawdopodobieństwem podobny do prekursora właściwościami chemicznymi. A więc wiele mutacji punktowych ma szansę wprowadzić do zmutowanego białka aminokwas podobny, a nie skrajnie odmienny, co zmniejsza ryzyko konsekwencji takich mutacji. Oto rozwiązanie problemu 9.
SAMOORGANIZACJA MATERII
Praca detektywów była ciężka, a wyniki które uzyskali - ważne. Była to jednak głównie próba odpowiedzenia na pytanie: jak powstały najważniejsze składniki obecnych żywych komórek i jaka była ich kolejność pojawiania się na ewolucyjnej scenie? Istnieją jednak jeszcze inne, bardzo ważne kwestie: jak organizowały się te składniki by utworzyć prakomórkę? Jaka była kolejność integracji metabolizmu, błony komórkowej i informacji genetycznej?
Tibor Ganti w swej książce: „Podstawy życia” przedstawił abstrakcyjny model minimalnego systemu żywego. Nazwa: minimalny pochodzi stąd, że do tego, aby system wykazywał nową cechę - życie potrzebne są wszystkie jego składniki. Brak jakiegokolwiek z nich oznacza śmierć.
Ganti nazwał ów system chemotonem. Nie musiał on w rzeczywistości istnieć, ale spełnia już wszystkie kryteria życia przyjęte przez autora. I spróbujmy teraz oprzeć się na tym pomyśle i spróbować dopasować go do danych, z którymi zapoznaliśmy się.
Zanim jednak do tego przystąpimy spójrzmy na konkretne dane z dziedziny biochemii, które będą nam w tym pomocne.
U roślin i mikroorganizmów występuje specyficzny cykl przemian biochemicznych, zwany cyklem glioksalowym. Pozwala on im na syntetyzowanie cukrów z tłuszczów. Jest on szczególnie istotny dla roślin oleistych takich jak soja, rzepak, słonecznik. Ich nasiona zawierają tłuszcz jako materiał zapasowy. Rozwijający się zarodek potrzebuje również cukrów, które uzyskuje właśnie przez przemianę tłuszczów w tym cyklu. Zwierzęta, w tym także ludzie, nie mają tej zdolności. Potrafią przeprowadzać tylko reakcję odwrotną: cukier → tłuszcz. Wiedzą o tym doskonale amatorzy różnych łakoci.
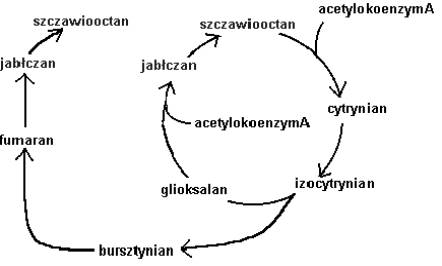
Rys.6. Schemat cyklu glioksalowego. Strzałka oznacza reakcję katalizowaną przez określony enzym.
Ci z nich, którzy nie chcą pogodzić się z powiedzeniem: „ Cierp ciało jeśliś chciało” muszą starać się spalić pewien nadmiar tkanki tłuszczowej (na pewno nie taki wielki...). Wymaga to wysiłku fizycznego i z pewnością nie robi się wtedy nikomu słodko na duszy właśnie dlatego, że tłuszcz może u człowieka zamienić się tylko w CO2 i H2O (pocimy się), a nie na przykład - we fruktozę.
Powyższy cykl reakcji możemy traktować jako katalizator, czyli związek lub ich grupa które pomagają w zajściu reakcji, lecz same w jej wyniku się nie zmieniają. Widzimy że po przejściu substratu - acetylokoenzymu A w główny produkt - bursztynian cykl powraca do punktu wyjścia . Koło się zamyka.
Reakcja jest zbyt złożona, aby przeprowadzić ją za pomocą jednego enzymu, stąd cały ich zespół uzupełniony jeszcze o organiczne związki niskocząsteczkowe. Gdyby był to ciąg, a nie cykl reakcji, mógłby również katalizować złożone przemiany, lecz nie byłoby możliwości powrotu do stanu początkowego (zob.rys. 7).
Tak naprawdę nie byłby to więc katalizator (te nie zmieniają się wskutek reakcji, których przebieg ułatwiają). Komórka stosująca ciąg musiałaby dostarczać nie tylko substratu S, lecz także związku X. Otrzymywałaby również produkt uboczny K, który musiałaby usuwać. W cyklu na rysunku 7 związek X regeneruje się ze związku K.
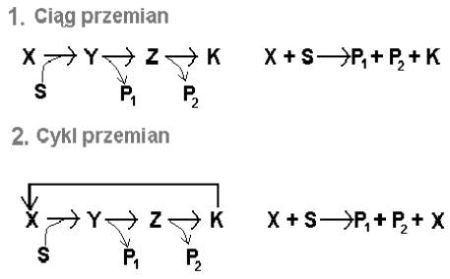
Rys.7. Porównanie cyklu i ciągu przemian.
Załóżmy teraz, że jednym z produktów P w cyklu przemian jest związek X. Mielibyśmy wtedy właśnie taką sytuację jak w cyklu glioksalowym. Schematycznie przedstawia ją rys. 8. W tym szczególnym przypadku związkiem X jest jabłczan.
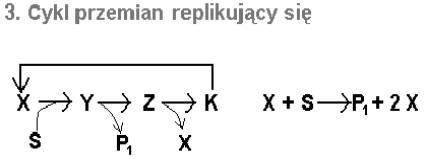
Rys.8. Schemat cyklu przemian zdolnego do powielania samego siebie.
Cykl po jednej rundzie wytwarza jedną cząsteczkę jabłczanu zużywając 2 cząsteczki substratu - acetylokoenzymu A (AcCoA). Powstały jabłczan może zainicjować kolejną przemianę z zaangażowaniem dwóch AcCoA i znów wytworzyć dodatkową cząsteczkę jabłczanu, itd.
Ogólnie więc: w n cyklach 2n cząsteczek AcCoA wytwarza n nowych cząsteczek jabłczanu, które mogą zapoczątkować n kolejnych cykli z pochłonięciem 2n AcCoA.
Czyli, im więcej AcCoA zostaje pochłonięte, tym więcej powstanie cząsteczek do jego dalszego pochłonięcia. Zjawisko, dzięki któremu jakiś proces ma zdolność do nasilania własnego przebiegu nazywamy sprzężeniem zwrotnym dodatnim.
Nie może on jednak trwać w nieskończoność. Liczba dostępnych AcCoA jest przecież ograniczona i zależy od natężenia rozpadu kwasów tłuszczowych, którego acetylokoenzym jest końcowym produktem.
Postarajmy się teraz dopasować cykl pentozofosforanów do schematu chemotonu.
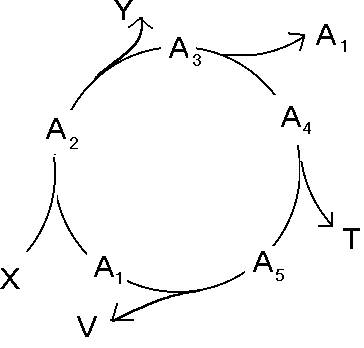
Schemat chemotonu
Oto jego zapis sumaryczny: 6 glukoz + 12 NADP → 12 NADPH2 + 5 glukoz + 6CO2
Substrat X to glukoza, produkt Y to CO2 i NADPH2. Jednym ze związków pośrednich tej reakcji jest rybozo-5fosforan, monomer kwasu polifosforybozowego, czyli V.
Nie jest to co prawda produkt, lecz nie jest również powiedziane, że w komórce, w której pracuje wiele cykli, wszystkie muszą się zamykać. W terminologii biochemicznej mówi się, że cykle mogą pracować w różnych trybach operacyjnych.
Część z nich może dostarczać związków potrzebnych komórce nawet jeśli są one częścią cyklu! (zob. rys.9). Związki te nie mogą tylko zostać wyczerpane całkowicie, gdyż wtedy wszystkie cykle załamałyby się. Zauważmy jednak, że cykl pentozofosforanów nie ma zdolności autoreplikowania się. Liczba glukoz zmniejsza się po przebiegu reakcji, zamiast zwiększać się.
Problemem pozostaje również istnienie związku symbolizowanego przez T. Współcześnie błona komórkowa budowana jest głównie przez fosfolipidy, których synteza jest skomplikowana i nie ma nic bezpośrednio wspólnego z cyklem pentozofosforanów. Jak jednak pamiętamy, koacerwaty tworzone były m.in. z polisacharydów, więc być może jakieś polimery cukrów uczestniczących w tym cyklu wchodziły w skład pierwotnej plazmalemmy.
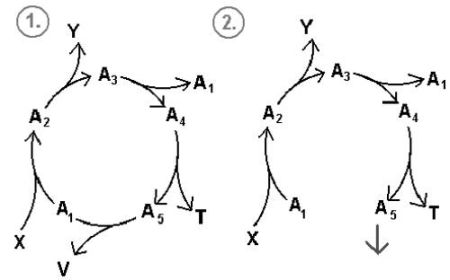
Rys.9. Dwa tryby operacyjne cyklu chemicznego.
1. Schemat klasycznego cyklu.
2. Schemat cyklu, którego jeden ze składników pełni rolę produktu. Cykl zamienia się w ciąg i może istnieć w zależności od dostaw związku A4, A3, A1, A2.
Wiemy, że cykl pentoz występuje w całym świecie ożywionym. Dane te i częściowe dopasowanie do schematu chemotonu sugerują, że pierwotny cykl metaboliczny mógł być podobny do cyklu pentoz. Dostawa cukrów do reakcji była łatwa, bo powstawały one obficie w reakcji formozowej (problem 3).
Potem cykl ten opłaszczył się otoczką polisacharydową, a dalej mógł powstać polimer informacyjny (kwas polifosforybozowy z rysunku 3 lub nawet łańcuch złożony z samych ryboz), służący pierwotnie jako materiał zapasowy. Pierwotny metabolizm był początkowo prawie na pewno oparty na cukrach, z czasem wytworzyła się błona komórkowa, a dalej polimer informacyjny.
Możliwe, że po powstaniu błon lipidowych rozwinęła się chemosynteza, czyli synteza związków potrzebnych organizmowi z udziałem energii zawartych w wysokoenergetycznych związkach nieorganicznych, takich jak metan lub siarkowodór. Związki te łatwo przechodzą przez błony lipidowe. Chemosynteza nie mogła powstać od razu, bo wymaga skomplikowanego zestawu enzymów.
Przypuszczalnie najpierw prakomórki bazowały na cukrach, łatwo powstających w środowisku zewnętrznym z formaldehydu, i mogących w izolowanych zbiornikach osiągnąć duże stężenia.
PODZIAŁ KOMÓRKI
a) Podział genetycznie niekontrolowany (część twórcza)
Z zasad geometrii wiemy, że powierzchnia (S) jest proporcjonalna i wzrasta proporcjonalnie do kwadratu wymiarów liniowych (a). Objętość (V) natomiast jest w taki sposób powiązana z sześcianem wymiarów liniowych. Kiedy zwiększymy dwukrotnie długość boku kwadratu, jego powierzchnia wzrośnie 22 = 4 razy. Podwojenie długości boków sześcianu sprawi, że jego objętość wzrośnie 23 = 8 razy. Możemy więc napisać:
S ~ a^2 , V ~ a^3
W jakiej relacji pozostają jednak S i V ? Przekształćmy powyższe wzory:
a ~ V^1/3 → S ~ V^2/3
Interesujące będzie zbadanie jak ze wzrostem objętości zmienia się stosunek: S/V. Podzielmy wzór:
S ~ V^2/3 przez V
S/V ~ V^-1/3 → S/V = m V^-1/3 , m - wsp.proporcjonalności
Jedną z takich funkcji przedstawia wykres 1:
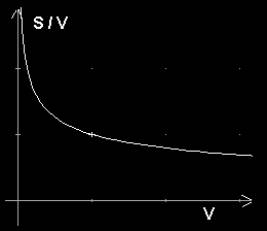
Wykres 1. Funkcja S/V = m V^-1/3 dla m = 1.
Jest to funkcja malejąca. Wraz ze wzrostem argumentu V maleje wartość S/V.
Kiedy komórka (którą przedstawimy abstrakcyjnie za pomocą kuli) dzieli się, to powstają dwie komórki potomne o objętości v = 0,5 V.
Wzory na ich objętość wyglądają następująco:
V = 4/3 π R^3 komórka macierzysta
v = 4/3 π r^3 komórka potomna
Można łatwo obliczyć, że promienie tych kul pozostają w relacji: R = r 2^1/3.
Jak wynika z wykresu 1, wraz ze wzrostem objętości maleje stosunek powierzchni do objętości, z czego wynika, że powierzchnia rośnie wolniej niż objętość.
Komórka, mająca 2 razy większą objętość, nie będzie miała 2 razy większej powierzchni. Wzory naszych komórek na powierzchnię wyglądają następująco:
P = 4 π R^2 = 4 π (r 2^1/3 )^2 = 4 π r^2 2^2/3 komórka macierzysta
p = 4 π r^2 komórka potomna
Można zauważyć, że P < 2p, bo 2^2/3 < 2
Zróbmy teraz realne założenie, że prymitywny składnik błony komórkowej T produkowany jest w cytoplazmie, całą objętością komórki. Z tego wniosek, że powierzchnia chemotonu będzie wzrastać proporcjonalnie do jego objętości według wzoru:
S ~ V, S = kV, k - wsp. proporcjonalności
Powierzchnia chemotonu ma konkretną postać:
Sch = k 4/3 π R^3
Sch = k 4/3 π R^3 = k 4/3 π r^3 (2^1/3)^3 = 2 k 4/3 π r^3
Postawmy teraz warunek podziału: Sch = 2p, czyli:
2 k 4/3 π r^3 = 8 π r^2 → r = 3/k , R = 3/k 2^1/3
Gdy prakomórka osiągnie promień R równy 3/k 2^1/3, będzie miała 2 razy większą objętość i 2 razy więcej materiału na powierzchnię, niż komórka potomna. Proces podziału będzie więc wtedy możliwy. I nigdzie nie musi być zapisywany!
b) Rozdział materiału genetycznego
Załóżmy że na początku istnieją tylko 4 rodzaje cząstek RNA i są one dla komórki niezbędne. Niekoniecznie replikują się one w tym samym tempie. Komórka dzieli się i są one losowo rozdzielane do komórek potomnych. Jakie są tu zagrożenia? Liczba kopii RNA, będących niezależnymi cząsteczkami nie jest zsynchronizowana. Niektóre mogą dzielić się szybciej – inne wyraźnie wolniej. Poza tym z przyczyn czysto losowych komórka potomna może nie otrzymać wszystkich czterech rodzajów cząstek.
Problemy te rozwiązuje związanie ze sobą wszystkich czterech cząstek na jedną, czterogenową nić. Wtedy to na pewno wszystkie cztery geny trafią do komórki potomnej i będą replikowane wolniej (są na dłuższej cząstce), ale w takiej samej liczbie kopii. Te korzyści zwyciężyły (problem 10). I dlatego powstały chromosomy – długie nici DNA z zapisanymi w nich sekwencjami wielu genów.
EWOLUCJA EGZONÓW I INTRONÓW
Introny dzielące sekwencje genów na kawałki, występują powszechnie u Eucaryota. W przypadku Procaryota mają je tylko archebakterie.
Istnieją dwie hipotezy, wyjaśniające istnienie przerywanych genów u Eucaryota:
1. Wspólny przodek Eucaryota i Procaryota miał już geny nieciągłe. Wszystkie organizmy prokariotyczne utraciły introny w wyniku presji selekcyjnej na minimalizację zbędnej informacji.
2. Wspólny przodek Eucaryota i Procaryota miał geny ciągłe. Eucaryota nabyły sekwencji intronowych w wyniku insercji.
Hipotezy powyższe nie wykluczają się całkowicie. Można przecież wyobrazić sobie, że wspólny przodek eukariontów i prokariontów posiadał introny, które następnie zostały przez wszystkie bakterie i sinice utracone, natomiast Eucaryota zyskiwały dodatkowe kolejne introny przez insercje.
Właśnie ta synteza dwóch powyższych hipotez wydaje się najlepiej potwierdzać w rzeczywistości, gdyż istnieją doniesienia o sporadycznej obecności u eukariontów intronów pochodzenia insercyjnego. Jest to jednak zjawisko rzadkie i specyficzne dla pojedynczych gatunków. Możemy zatem wnioskować, że jest ono wtórną nakładką na introny dziedziczone po przodkach, istniejące w pierwotnych genach, podobnie jak obecność u kilku gatunków nietypowych kodonów jest nakładką na uniwersalny kod genetyczny.
Pozostaje nam jeszcze wyjaśnić przyczynę silnej presji selekcyjnej na redukcję intronów w ewolucji Procaryota. Przedtem jednak zapoznajmy się z definicją 2 ogólnie wyodrębnianych strategii przetrwania organizmów:
1. Strategia r - Prawdopodobieństwo przeżycia dla pojedynczego potomka niewielkie. Organizm wydaje dużą liczbę potomstwa, aby przeżył niewielki jego procent.
2. Strategia K - Prawdopodobieństwo przeżycia pojedynczego potomka duże ze względu na opiekę rodzicielską. Organizm wydaje niewielką liczbę potomstwa i przeżywa duży jego procent.
Powyższe przypadki są oczywiście skrajne. Gatunki w przyrodzie realizują z reguły strategie pośrednie. Procaryota są jednak bardzo blisko strategii r, Eucaryota natomiast stosują strategie różne w zależności od gatunku z podkreśleniem, że duże wyspecjalizowane organizmy z tej grupy (np. ptaki, ssaki) stosują strategię K. Trzeba tu też nadmienić, że od pojęć strategii r i K ze względu na ich skrajność ostatnimi czasy odchodzi się.
Istnieje związek pomiędzy strategią r u bakterii i sinic, a brakiem w ich genach intronów. W ewolucji wygrały bowiem te z nich, które były w stanie wydać największą liczbę potomstwa. U prokariontów rozmnażanie odbywa się przez podział komórki. Duża liczba podziałów w jednostce czasu znaczyła więc dużą liczba potomków. Zanim komórka się podzieli, musi zreplikować cały swój materiał genetyczny, a żeby zrobić to jak najszybciej, musi on być jak najkrótszy. Dlatego genomy prokariotycznych zawierają informacje na temat, pozbawione wszelkich wtrętów. Organizmy z takimi oszczędnymi genomami rozprzestrzeniają się najszybciej dominując i wypierając pozostałe. Zmienność jest ważna, aby istniała możliwość przetrwania niektórych komórek, gdy zmienia się środowisko. Jej źródłem u Procaryota są przede wszystkim mutacje.
Źródłem zmienności u Eukaryota są natomiast głównie rekombinacje, czyli tworzenie nowych zestawień istniejących już alleli lub tworzących je egzonów.
Spójrzmy na proces rekombinacji, który odbywa się wewnątrz jakichś 2 alleli:

Obecność intronów zapewnia tu:
1. Zwiększone prawdopodobieństwo rekombinacji między egzonami, bo wymiana egzonów nastąpi zawsze przy przerwaniu jakiegokolwiek z wielu wiązań w intronie oddzielającym je.
2. Brak wymogu precyzji przy rekombinacji. W części 1 powyższego rysunku istnieje nie 5 lecz aż 25 możliwości zajścia rekombinacji, gdyż może nastąpić przerwanie na intronie wiązań nieodpowiadających sobie. Gdyby intron nie istniał (co obrazuje sytuacja w części 2 rysunku) byłaby tylko jedna możliwość rekombinacji, musiałaby więc ona być precyzyjna, by nie zniszczyć zapisu w egzonie, a jeśli taką by nie była – spowodowałaby mutację.
W przypadku strategii K, ilość materiału genetycznego w komórce nie jest tak istotna, gdyż tu nie liczy się krótki czas trwania pokolenia. Wręcz przeciwnie - długość życia osobnika jest bardzo długa w porównaniu z okresem międzypodziałowym u bakterii i sinic (np. u bakterii Escherichia coli - 20 minut).
Tu liczy się przystosowanie pojedynczych osobników, ponieważ prawdopodobieństwo ich przeżycia musi być wysokie, skoro rodzi się ich niewiele. Tak więc ważna jest możliwość tworzenia kombinacji nowych alleli, gdyż te mogą okazać się plastyczniejsze („zawsze jakaś dostosuje się”).
W przypadku eukariontów z wyraźną strategią r, jakimi są np. jednokomórkowe drożdże, obserwujemy znaczną redukcję sekwencji intronowych w ich genach.
REKONSTRUKCJA WYDARZEŃ
Po zaznajomieniu się z wynikami śledztwa, przedstawionymi przez wszystkich detektywów, możemy pokusić się o próbę hipotetycznej rekonstrukcji wydarzeń:
1. Oceany przed 3,8 mld lat obfitowały w fosforany i formaldehyd. Podobnie mogło być z formaldehydem spadającym do takich lądowych kałuż wraz z deszczem. Z czasem formaldehyd kondensował z utworzeniem monosacharydów. One zaś mogły wiązać grupy fosforanowe, by zamienić się w fosfocukry (reakcje katalizowane np. przez piryt).
2. Fosfocukry formują prymitywne cykle, powstaje pierwszy cykl autoreplikujący się. Reakcje zachodzą bardzo powoli, gdyż nie są katalizowane.
3. Powstaje otoczka komórkowa oparta na polisacharydach (?).
4. Tworzą się pierwsze polimery informacyjne, oparte na kwasie polifosforybozowym.
5. Do polimeru podłączone zostają reszty aminokwasów, produkowanych w atmosferze gazów redukujących, wskutek wyładowań atmosferycznych. Powstają polimery o zdolnościach katalitycznych.
6. Polimery katalizują reakcje metaboliczne i dołączanie randomiczne HCHO, NH3, CO2 na innych polimerach, co prowadzi do powstania zasad azotowych i w konsekwencji RNA.
7. Powstają pierwsze 100-200 nukleotydowe rybozymy, które mogą katalizować reakcje metaboliczne, replikować się z udziałem interkalujących związków porfirynowych i/lub dzięki wzajemnym wspieraniu się (hipercykle). Na początku w ich skład wchodzi przypuszczalnie tylko guanina i cytozyna.
8. Powstają pierwsze cząstki tRNA, do których podłączają się stopniowo aminokwasy (najpierw glicyna, alanina, walina, kwas asparaginowy). Następuje prymitywna synteza białek na matrycy prymitywnego mRNA.
Skomplikowane kompleksy rybozymów zaczynają wspomagać biosyntezę białek. Powstają pierwsze rybosomy.
9. Aminokwasy „tylko L” i cukry tylko „D” utrwalają się w przodkach obecnych organizmów. Przypuszczalnie odpowiada za to wspomaganie przez reakcje, w których brały udział te związki przez powierzchnie faworyzujące aminokwasy L i cukry D. Nie należy też lekceważyć teoretycznych rozważań fizyków, którzy mówią, że rozpad beta jest w stanie zdestabilizować bardziej aminokwasy D niż L. Jak obliczono, do prawie pewnej wygranej aminokwasów L w wyniku tego nieco selektywnego procesu, potrzebne byłoby 15 000 lat. Hipotezę tę trudno więc przetestować w laboratorium (problem 7).
10. W wyniku powstania białek, RNA zostaje zamieniony na stabilny, dwuniciowy DNA. Następuje rozdział funkcji pomiędzy białka (enzymy, składniki budulcowe) i DNA (zapis informacji genetycznej). Białka zaczynają odczytywać informację z DNA, syntetyzując na podstawie jego sekwencji prymitywne mRNA. Powstaje transkrypcja.
WYNIK: Organizm żywy, jednokomórkowy, uzyskujący energię ze źródeł zewnętrznych, mający swoją błonę komórkową, materiał genetyczny, maszynerię enzymatyczną, zdolność zapisu genetycznego i biosyntezy białek.
ZAKOŃCZENIE
Czy zauważył Czytelnik jak często w tym rozdziale pojawiły się pochodne wyrazu „może”? Wywody są często mgliste, a cała ta rekonstrukcja jest jednym, wielkim domniemaniem. Są to tylko próby odpowiedzi na postawione trudne biogenetyczne problemy i propozycje rozwiązań. Niewielu rzeczy tutaj tak naprawdę jesteśmy pewni. Ale samo postawienie problemów jest już osiągnięciem.
Tibor Ganti - zbudował abstrakcyjny model systemu żywego. Można więc powiedzieć, że wyznaczył pewne ramy, granice zbioru życia. Nie uzyskujemy jednak od niego konkretnych informacji o tym, jak skonstruować sztucznie komórkę żywą w labolatorium, najlepiej identyczną z tą pierwotną, sprzed 3 mld lat. Co dokładnie i kolejno robić ?
Mówiąc językiem detektywów, dzięki niemu wiemy tylko do jakiej grupy mógł należeć „przestępca”.
Badając ślady, jakie pozostawił w obecnie istniejących komórkach (cykl pentoz oparty na fosfocukrach, rybozymy, biosynteza puryn i pirymidyn, uniwersalny kod genetyczny, te same kwasy nukleinowe RNA i DNA, jedność aminokwasów) możemy starać się szkicować jego coraz bardziej szczegółowe portrety pamięciowe, a właściwie „śladowe”.
To wszystko co możemy zrobić. Na zawsze nieznane pozostanie miejsce i czas powstania życia, nigdy nie dowiemy się konkretnie jak wyglądała prakomórka, jaki dokładnie miała metabolizm i materiał genetyczny. A zatem samego „przestępcy” nie złapiemy już nigdy. Czy jednak musimy go znać...?
Maciej Panczykowski
Artykuł pochodzi z Wortalu Nauk Przyrodniczych Autora: http://mpancz.webpark.pl
Dziękujemy za zgodę na publikację tekstu.