Jak powstało życie?
Nagromadzenie danych to nie jest jeszcze nauka. Galileusz .:::::.
Cykl po jednej rundzie wytwarza jedną cząsteczkę jabłczanu zużywając 2 cząsteczki substratu - acetylokoenzymu A (AcCoA). Powstały jabłczan może zainicjować kolejną przemianę z zaangażowaniem dwóch AcCoA i znów wytworzyć dodatkową cząsteczkę jabłczanu, itd.
Ogólnie więc: w n cyklach 2n cząsteczek AcCoA wytwarza n nowych cząsteczek jabłczanu, które mogą zapoczątkować n kolejnych cykli z pochłonięciem 2n AcCoA.
Czyli, im więcej AcCoA zostaje pochłonięte, tym więcej powstanie cząsteczek do jego dalszego pochłonięcia. Zjawisko, dzięki któremu jakiś proces ma zdolność do nasilania własnego przebiegu nazywamy sprzężeniem zwrotnym dodatnim.
Nie może on jednak trwać w nieskończoność. Liczba dostępnych AcCoA jest przecież ograniczona i zależy od natężenia rozpadu kwasów tłuszczowych, którego acetylokoenzym jest końcowym produktem.
Postarajmy się teraz dopasować cykl pentozofosforanów do schematu chemotonu.
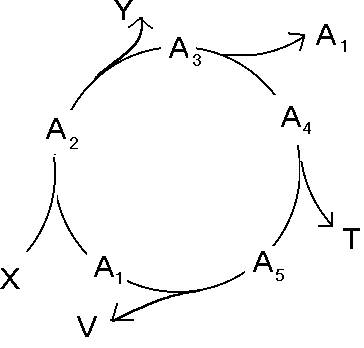
Schemat chemotonu
Oto jego zapis sumaryczny: 6 glukoz + 12 NADP → 12 NADPH2 + 5 glukoz + 6CO2
Substrat X to glukoza, produkt Y to CO2 i NADPH2. Jednym ze związków pośrednich tej reakcji jest rybozo-5fosforan, monomer kwasu polifosforybozowego, czyli V.
Nie jest to co prawda produkt, lecz nie jest również powiedziane, że w komórce, w której pracuje wiele cykli, wszystkie muszą się zamykać. W terminologii biochemicznej mówi się, że cykle mogą pracować w różnych trybach operacyjnych.
Część z nich może dostarczać związków potrzebnych komórce nawet jeśli są one częścią cyklu! (zob. rys.9). Związki te nie mogą tylko zostać wyczerpane całkowicie, gdyż wtedy wszystkie cykle załamałyby się. Zauważmy jednak, że cykl pentozofosforanów nie ma zdolności autoreplikowania się. Liczba glukoz zmniejsza się po przebiegu reakcji, zamiast zwiększać się.
Problemem pozostaje również istnienie związku symbolizowanego przez T. Współcześnie błona komórkowa budowana jest głównie przez fosfolipidy, których synteza jest skomplikowana i nie ma nic bezpośrednio wspólnego z cyklem pentozofosforanów. Jak jednak pamiętamy, koacerwaty tworzone były m.in. z polisacharydów, więc być może jakieś polimery cukrów uczestniczących w tym cyklu wchodziły w skład pierwotnej plazmalemmy.
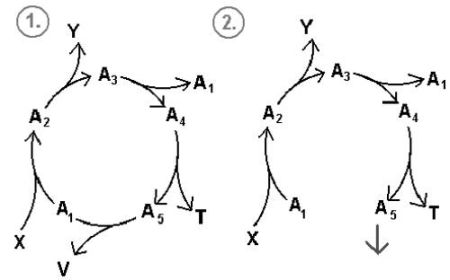
Rys.9. Dwa tryby operacyjne cyklu chemicznego.
1. Schemat klasycznego cyklu.
2. Schemat cyklu, którego jeden ze składników pełni rolę produktu. Cykl zamienia się w ciąg i może istnieć w zależności od dostaw związku A4, A3, A1, A2.
Wiemy, że cykl pentoz występuje w całym świecie ożywionym. Dane te i częściowe dopasowanie do schematu chemotonu sugerują, że pierwotny cykl metaboliczny mógł być podobny do cyklu pentoz. Dostawa cukrów do reakcji była łatwa, bo powstawały one obficie w reakcji formozowej (problem 3).
Potem cykl ten opłaszczył się otoczką polisacharydową, a dalej mógł powstać polimer informacyjny (kwas polifosforybozowy z rysunku 3 lub nawet łańcuch złożony z samych ryboz), służący pierwotnie jako materiał zapasowy. Pierwotny metabolizm był początkowo prawie na pewno oparty na cukrach, z czasem wytworzyła się błona komórkowa, a dalej polimer informacyjny.
Możliwe, że po powstaniu błon lipidowych rozwinęła się chemosynteza, czyli synteza związków potrzebnych organizmowi z udziałem energii zawartych w wysokoenergetycznych związkach nieorganicznych, takich jak metan lub siarkowodór. Związki te łatwo przechodzą przez błony lipidowe. Chemosynteza nie mogła powstać od razu, bo wymaga skomplikowanego zestawu enzymów.
Przypuszczalnie najpierw prakomórki bazowały na cukrach, łatwo powstających w środowisku zewnętrznym z formaldehydu, i mogących w izolowanych zbiornikach osiągnąć duże stężenia.
aktualna ocena | |
głosujących | |
 Ocena |
bardzo słabe |
słabe |
średnie |
dobre |
super |
Ocena |
bardzo słabe |
słabe |
średnie |
dobre |
super |

















W czasie ciąży można bezpiecznie prowadzić leczenie onkologiczne
Choroba nowotworowa nie jest wskazaniem do przedwczesnego rozwiązania ciąży.












