Jak powstało życie?
Nagromadzenie danych to nie jest jeszcze nauka. Galileusz .:::::.
Jak jednak powstały cząsteczki RNA z łańcucha polifosforybozowego?
Stawiam tutaj hipotezę o powstaniu zasad azotowych RNA, wykorzystując wiedzę biochemiczną o współcześnie istniejącym metabolizmie.
Biosynteza puryn odbywa się w komórkach z udziałem glicyny, formaldehydu i amoniaku (specyficznie związanych). Biosynteza pirymidyn wymaga natomiast kwasu asparaginowego, amoniaku i dwutlenku węgla. Wszystkie te substancje występowały już w prakałuży i praatmosferze. Zdumiewająca zgodność !
Związek z rys.2 mógł więc mieć najpierw doczepiany w miejsce grup 1’OH amoniak, potem aminokwasy, co zwiększało jego zdolności katalityczne. Teraz mógł katalizować dalsze dobudowywanie reszt HCHO i NH3 na innych łańcuchach, aż do całkowitego zsyntetyzowania zasad azotowych (zob. rys.3)
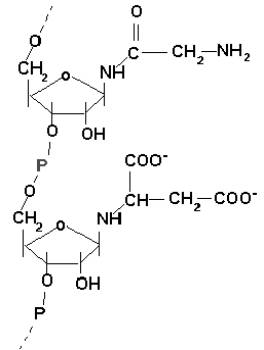
Rys.3. Wzór strukturalny domniemanego związku po dołączeniu do kwasu polifosforybozowego reszt amoniaku, glicyny i kw.asparaginowego.
c) Detektyw Eigen
Manfred Eigen skupił swą uwagę na cząsteczkach zdolnych do replikacji. Jego eksperyment z replikazą RNA, pochodzącą z wirusa o nazwie Qbeta pokazał, że replikowane przez nią cząsteczki RNA „walczą” o byt, a zmagania te zawsze doprowadzają do wyselekcjonowania cząsteczki, która replikuje się najwydajniej w środowisku, które stanowi określone stężenie soli i temperatura.
Eksperyment jest powtarzalny. Problemem jest fakt, że replikaza jest białkiem, które miało przecież w ewolucji powstać później. Orgel wykazał jednak, że cząsteczki RNA są zdolne do autoreplikacji. Trzeba im tylko dostarczyć rybonukleotydów i kationów cynku.
J. Maynard Smith i E. Szathmary udowadniają, że przy założeniu, że szybkość replikacji cząsteczek RNA na dwie wolne nici jest znacznie większa niż szybkość dysocjacji podwójnej helisy RNA, powstałej w replikacji, możliwa jest stabilna koegzystencja różnych cząstek RNA. Charakteryzuje je wtedy wzrost paraboliczny. Liczy się nie tylko szybkość produkcji kopii, ale także szybkość oddysocjowywania wolnych, pojedynczych nici do dalszej syntezy.
Wzrost opisuje równanie: dA/dt = kA^p , gdzie p<1. W takim przypadku cząsteczki RNA nie „wygryzają się” wzajemnie w taki sposób, że zostanie tylko jedna.
Wzrost, który zależy tylko od szybkości produkcji kopii opisywany wzorem:
dA/dt = kA^p, (p=1) to wzrost ekspotencjalny charakterystyczny dla teoretycznego, nieograniczonego wzrostu kolonii komórek. Tam czynnika dysocjacji nie ma, bo one nie są ze sobą „sklejone” wiązaniami chemicznymi.
Cząsteczki RNA potrafią mieć własności katalityczne. Może więc potrafią wzajemnie wspomagać się w replikacji? Myśl ta naprowadziła Manfreda Eigena na ideę hipercykli (zob. rys.4).
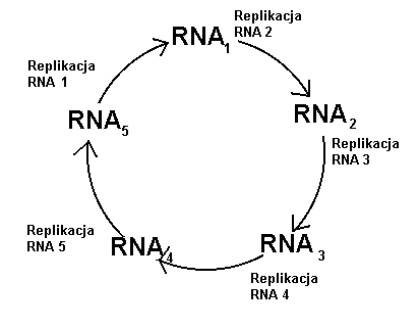
Rys.4. Schemat hipercyklu.
aktualna ocena | |
głosujących | |
 Ocena |
bardzo słabe |
słabe |
średnie |
dobre |
super |
Ocena |
bardzo słabe |
słabe |
średnie |
dobre |
super |

















W czasie ciąży można bezpiecznie prowadzić leczenie onkologiczne
Choroba nowotworowa nie jest wskazaniem do przedwczesnego rozwiązania ciąży.












