Jak powstało życie?
Nagromadzenie danych to nie jest jeszcze nauka. Galileusz .:::::.
EWOLUCJA EGZONÓW I INTRONÓW
Introny dzielące sekwencje genów na kawałki, występują powszechnie u Eucaryota. W przypadku Procaryota mają je tylko archebakterie.
Istnieją dwie hipotezy, wyjaśniające istnienie przerywanych genów u Eucaryota:
1. Wspólny przodek Eucaryota i Procaryota miał już geny nieciągłe. Wszystkie organizmy prokariotyczne utraciły introny w wyniku presji selekcyjnej na minimalizację zbędnej informacji.
2. Wspólny przodek Eucaryota i Procaryota miał geny ciągłe. Eucaryota nabyły sekwencji intronowych w wyniku insercji.
Hipotezy powyższe nie wykluczają się całkowicie. Można przecież wyobrazić sobie, że wspólny przodek eukariontów i prokariontów posiadał introny, które następnie zostały przez wszystkie bakterie i sinice utracone, natomiast Eucaryota zyskiwały dodatkowe kolejne introny przez insercje.
Właśnie ta synteza dwóch powyższych hipotez wydaje się najlepiej potwierdzać w rzeczywistości, gdyż istnieją doniesienia o sporadycznej obecności u eukariontów intronów pochodzenia insercyjnego. Jest to jednak zjawisko rzadkie i specyficzne dla pojedynczych gatunków. Możemy zatem wnioskować, że jest ono wtórną nakładką na introny dziedziczone po przodkach, istniejące w pierwotnych genach, podobnie jak obecność u kilku gatunków nietypowych kodonów jest nakładką na uniwersalny kod genetyczny.
Pozostaje nam jeszcze wyjaśnić przyczynę silnej presji selekcyjnej na redukcję intronów w ewolucji Procaryota. Przedtem jednak zapoznajmy się z definicją 2 ogólnie wyodrębnianych strategii przetrwania organizmów:
1. Strategia r - Prawdopodobieństwo przeżycia dla pojedynczego potomka niewielkie. Organizm wydaje dużą liczbę potomstwa, aby przeżył niewielki jego procent.
2. Strategia K - Prawdopodobieństwo przeżycia pojedynczego potomka duże ze względu na opiekę rodzicielską. Organizm wydaje niewielką liczbę potomstwa i przeżywa duży jego procent.
Powyższe przypadki są oczywiście skrajne. Gatunki w przyrodzie realizują z reguły strategie pośrednie. Procaryota są jednak bardzo blisko strategii r, Eucaryota natomiast stosują strategie różne w zależności od gatunku z podkreśleniem, że duże wyspecjalizowane organizmy z tej grupy (np. ptaki, ssaki) stosują strategię K. Trzeba tu też nadmienić, że od pojęć strategii r i K ze względu na ich skrajność ostatnimi czasy odchodzi się.
Istnieje związek pomiędzy strategią r u bakterii i sinic, a brakiem w ich genach intronów. W ewolucji wygrały bowiem te z nich, które były w stanie wydać największą liczbę potomstwa. U prokariontów rozmnażanie odbywa się przez podział komórki. Duża liczba podziałów w jednostce czasu znaczyła więc dużą liczba potomków. Zanim komórka się podzieli, musi zreplikować cały swój materiał genetyczny, a żeby zrobić to jak najszybciej, musi on być jak najkrótszy. Dlatego genomy prokariotycznych zawierają informacje na temat, pozbawione wszelkich wtrętów. Organizmy z takimi oszczędnymi genomami rozprzestrzeniają się najszybciej dominując i wypierając pozostałe. Zmienność jest ważna, aby istniała możliwość przetrwania niektórych komórek, gdy zmienia się środowisko. Jej źródłem u Procaryota są przede wszystkim mutacje.
Źródłem zmienności u Eukaryota są natomiast głównie rekombinacje, czyli tworzenie nowych zestawień istniejących już alleli lub tworzących je egzonów.
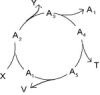
Spójrzmy na proces rekombinacji, który odbywa się wewnątrz jakichś 2 alleli:

Obecność intronów zapewnia tu:
1. Zwiększone prawdopodobieństwo rekombinacji między egzonami, bo wymiana egzonów nastąpi zawsze przy przerwaniu jakiegokolwiek z wielu wiązań w intronie oddzielającym je.
2. Brak wymogu precyzji przy rekombinacji. W części 1 powyższego rysunku istnieje nie 5 lecz aż 25 możliwości zajścia rekombinacji, gdyż może nastąpić przerwanie na intronie wiązań nieodpowiadających sobie. Gdyby intron nie istniał (co obrazuje sytuacja w części 2 rysunku) byłaby tylko jedna możliwość rekombinacji, musiałaby więc ona być precyzyjna, by nie zniszczyć zapisu w egzonie, a jeśli taką by nie była – spowodowałaby mutację.
W przypadku strategii K, ilość materiału genetycznego w komórce nie jest tak istotna, gdyż tu nie liczy się krótki czas trwania pokolenia. Wręcz przeciwnie - długość życia osobnika jest bardzo długa w porównaniu z okresem międzypodziałowym u bakterii i sinic (np. u bakterii Escherichia coli - 20 minut).
Tu liczy się przystosowanie pojedynczych osobników, ponieważ prawdopodobieństwo ich przeżycia musi być wysokie, skoro rodzi się ich niewiele. Tak więc ważna jest możliwość tworzenia kombinacji nowych alleli, gdyż te mogą okazać się plastyczniejsze („zawsze jakaś dostosuje się”).
W przypadku eukariontów z wyraźną strategią r, jakimi są np. jednokomórkowe drożdże, obserwujemy znaczną redukcję sekwencji intronowych w ich genach.
aktualna ocena | |
głosujących | |
 Ocena |
bardzo słabe |
słabe |
średnie |
dobre |
super |
Ocena |
bardzo słabe |
słabe |
średnie |
dobre |
super |

















Na Marsie odkryto ślady węglanów
Odkrycie przemawia za większymi szansami na istnienie w przeszłości życia na Czerwonej Planecie.

Nietypowa planeta okrąża dwie gwiazdy na prostopadłej orbicie
To pierwsze tak silne dowody na istnienie planety na orbicie biegunowej układzie podwójnym.



Nowe wskazówki odnośnie do przeszłości Marsa
Wskazują one na wulkaniczną przeszłość planety, co ma ogromne znaczenie dla poszukiwań śladów życia.



Z przepastnych archiwów


Kiedy mrugają wyścigowi kierowcy?
Jedno mrugnięcie to dla nich 20 metrów, na których wiele może się wydarzyć.


Ekspert: muzyka słuchana w trakcie jazdy wpływa na nasze bezpieczeństwo
Istotny jest zarówno rodzaj utworów, jak i poziom decybeli w kabinie pojazdu.

